|
Mesure de la glycémie.
Source: https://en.wikipedia.org/wiki/Glucose_test |
Il n’existe que des preuves limitées concernant la
diminution des complications macrovasculaires et de la mortalité à la suite d’interventions
multifactorielles pour le contrôle de la glycémie, de la pression artérielle et
des lipides chez les patients atteints de diabète de type 2. Cependant, les
cibles thérapeutiques sûres et efficaces pour contrer ces facteurs de risque n’ont
pas été précisément déterminées dans le cadre de telles interventions.
Dans cette étude ouverte, multicentrique,
randomisée, à groupes parallèles, effectuées dans 81 sites de recherche clinique
situés au Japon, nous avons réparti de manière aléatoire (1:1) des patients
atteints de diabète de type 2 âgés de 45-69 ans hypertendus ou dyslipidémiques,
ou les deux, présentant des valeurs de HbA1c de 6.9% (52.0 mmol/mol)
ou plus, pour recevoir un traitement classique de contrôle de la glycémie,
tension artérielle et des lipides (valeurs ciblées : HbA1c<6.9%
[mmol/mol], tension artérielle <130/80 mm Hg, LDL cholesterol <120 mg/dL
[ou 100 mg/dL chez des patients présentant un historique de maladie
coronarienne]) ou une thérapie intensive
(HbA1c <6.2% [44.3 mmol/mol], pression artérielle <120/75 mm
Hg, LDL cholesterol <80 mg/Dl [ou 70 mg/dL chez des patients avec historique
de maladie coronarienne]). La randomisation était effectuée à l’aide d’une méthode
dynamique de séquence générée par ordinateur, stratifiée selon le sexe, l’âge,
HbA1c, et l’historique de maladie cardiovasculaire. À la fois les
patients et les investigateurs avaient accès au tableau de randomisation. Le
critère principal d’évaluation était l’occurrence
d’événements comme infarctus du myocarde, AVC, revascularisation (pontage
coronarien, angioplastie coronaire transluminale, endartériectomie coronarienne,
angioplastie cérébrale percutanée transluminale, et pose d’endoprothèse
coronarienne), et mortalité toutes causes confondues. L’analyse principale
était effectuée sur la population en intention de traiter. (…).
Entre le 16 juin 2006 et le 31 mars 2009, 2 542
patients éligibles ont été répartis de manière aléatoire pour recevoir un
thérapie intensive ou un traitement conventionnel (1271 sujets dans chaque
groupe) et ont été suivis sur une période médiane de 8.5 ans (Intervalle
Interquartile [IQR] 7.3-9.0). Deux patients du groupe thérapie intensive se
sont révélés inéligibles après la randomisation, et ont donc été exclus des
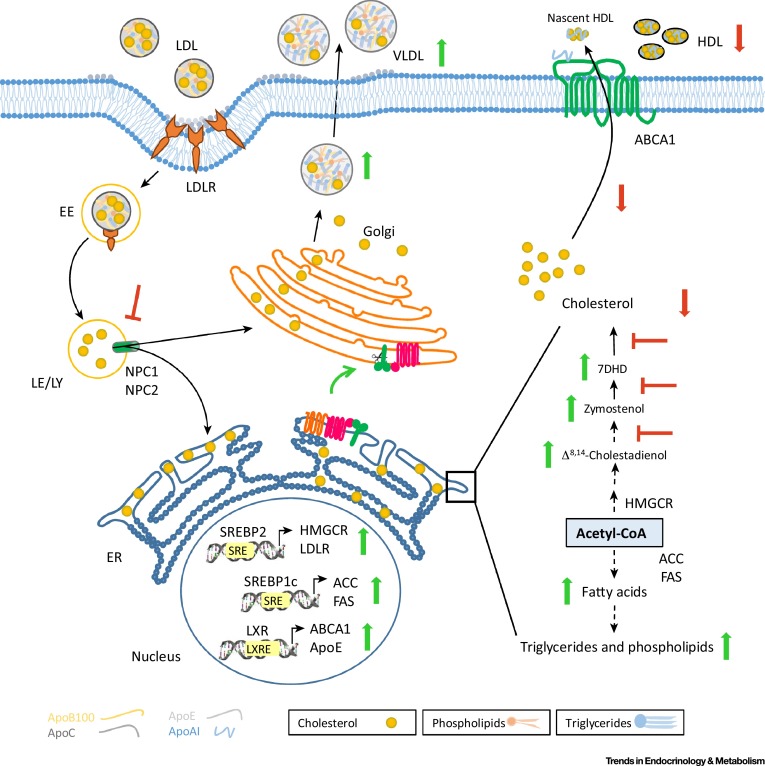
analyses. Au cours de la période d’intervention, les valeurs moyennes de HbA1c,
la pression artérielle systolique, la pression artérielle diastolique, et les
concentrations en LDL cholestérol étaient significativement plus basses dans le
groupe thérapie intensive que dans le groupe traitement conventionnel (6.8%
[51.0 mmol/mol] versus 7.2% [55.2
mmol/mol] ; 123 mm Hg versus 129
mm Hg ; 71 mm Hg versus 74 mm Hg ;
et 85 mg / dL versus 104 mg / dL,
respectivement ; p<0.0001
pour toutes les comparaisons). Le critère principal d’évaluation a été relevé
chez 109 patients du troupe thérapie intensive et chez 133 patients du groupe
thérapie conventionnelle (hazard ratio [HR] 0.81, Intervalle de Confiance [IC]
95% 0.63-1.04 ; p=0.094). (…)
Parmi tous les événements relevés, seuls les événements cérébrovasculaires
(AVC, angioplastie percutanée transluminale cérébrale, et pose d’endoprothèse
coronoarienne) étaient significativement moins fréquents dans le troupe
thérapie intensive (HR 0.42, 0.24-0.74 ; p=0.002).
Outre des événements d’hypoglycémie
non sévère (521 [41%] patients du groupe thérapie intensive versus 283 [22%]
dans le groupe thérapie conventionnelle, p<0.0001)
et d’œdème (193 [15%] versus 129 [10%], p=0.0001),
les fréquences des événements indésirables majeurs n’ont pas différé entre les
différents groupes.
Nos résultats ne montrent pas la pleine efficacité
d’une intervention multifactorielle intensifiée en comparaison des thérapies
standard actuellement en vigueur pour la prévention des différents événements
coronariens, cérébrovasculaires, et de la mortalité toutes causes confondues.
Quoi qu’il en soit, nos résultats suggèrent l’existence d’un bénéfice potentiel
d’une intervention intensifiée pour la prévention des événements
cérébrovasculaires chez les patients atteints de diabète de type 2. Prof
Kohijiro Ueki, MD, dans The Lancet Diabetes & Endocrinology, publication en
ligne en avant-première, 24 octobre 2017
Financement : Ministère
de la Santé, du Travail, et du Bien-Être du Japon, Asahi Kasei Pharma, Astellas
Pharma, AstraZeneca, Bayer, Boehringer Ingelheim, Bristol-Myers Squibb, Daiichi
Sankyo, Eli Lilly, GlaxoSmithKline, Kissei Pharmaceutical, Kowa Pharmaceutical,
Mitsubishi Tanabe Pharma, Mochida Pharmaceutical, MSD, Novartis Pharma, Novo
Nordisk, Ono Pharmaceutical, Pfizer, Sanwa Kagaku Kenkyusho, Shionogi, Sumitomo
Dainippon Pharma, Taisho Toyama Pharmaceutical, et Takeda.
Source : The Lancet Online / Traduction et adaptation : NZ